

תסיסה מאלולקטית

תסיסה מאלולקטית (MLF – malolactic fermentation) היא תסיסה המתרחשת בתהליכי תסיסה של תירוש המופק מזני ענבים המשמשים לייצור יין. הסובסטרט המשמש את התסיסה המאלולקטית הוא החומצה המאלית בעלת החומציות היותר גבוהה, המצויה באופן טבעי בתירוש ענבים והיא מומרת לחומצה לקטית בעלת החומציות היותר נמוכה בהשוואה לחומצה המאלית. תסיסה מאלולקטית מתבצעת לרוב כתסיסה משנית, זמן קצר לאחר סיום התסיסה ההתחלתית של סוכר הענבים והפיכתו לאתנול. היא עשויה לעיתים להתרחש במקביל לתסיסה האלכוהולית.
תהליך התסיסה הלאקטית נפוץ ברוב תהליכי ייצור היין האדום וגם עבור שימוש בזני ענבים לבנים, כמו שרדונה, המעניק טעם "חמאתי" שמקורו בהיווצרות דיאצטיל, (אנ') שהוא תוצר לוואי של התסיסה המאלולקטית.[1]
התהליך יכול להתרחש באופן טבעי על ידי הפלורה המצויה בתירוש. בייצור יין מסחרי, מתרחשת תסיסה מאלולקטית מבוקרת על ידי שימוש בחיידקים מתורבתים. הסטארטר הנפוץ ביותר לתסיסה מאלולקטית מכיל את החיידק Oenococcus oeni. התסיסה המאלולקטית מונעת התרבות של חיידקים ועובשים שמקורם בפלורה הטבעית של התירוש ויצירת טעמי "לוואי" בלתי רצויים ביין.
היסטוריה
מדען היין השווייצרי הרמן מולר היה מהראשונים שהעלו סברה שחיידקים הם הגורם להפחתת החומציות ביין. שליטה בתהליך התסיסה המאלולקטית היא פיתוח יחסי חדש. במשך מאות שנים, ייננים שמו לב ל"פעילות" שהתרחשה ביינותיהם המאוחסנים בחביות במהלך חודשי האביב החמים. בדומה לתסיסה אלכוהולית ראשונית, תופעה זו משחררת גז פחמן דו-חמצני הגורם לשינוי משמעותי של היין.[2]
בשנת 1837 היא תוארה כ"תסיסה שנייה" על ידי האונולוג הגרמני פרייהר פון באבו (אנ') וגרמה לעכירות מוגברת ביין. הוא עודד ייננים להגיב במהירות כשהבחינו פעילות זו, על ידי העברת היין לחבית חדשה, (אנ') הוספת גופרית דו-חמצנית לדיכוי הפעילות המיקרוביאלית, ולאחר מכן סבב נוסף של העברת היין לחביות חדשות והוספת מנה נוספת של גופרית-דו-חמצנית כדי לייצב את היין (אנ').[3] בשנת 1866, לואי פסטר, אחד מחלוצי המיקרוביולוגיה המודרנית, בודד את החיידקים הראשונים מיין וקבע שכל החיידקים הם גורמים קלקול היין. בעוד שפסטר שם לב לירידה בחומצה ביין בנוכחות חיידקי חומצת החלב, הוא לא קישר תהליך זה לצריכת חומצה מאלית על ידי החיידקים, אלא הניח שמדובר רק במשקעי מלחי הטרטרת (אנ').[2]
בשנת 1891, האונולוג השווייצרי הרמן מולר (אנ') העלה לראשונה תיאוריה שחיידקים עשויים להיות הגורם לתהליך, ובשנת 1913 הסביר את תהליך הפחת המוציות שנגרמת על ידי חיידק היין שנקראו בתחילה Baccilus gracilis והתברר לאחר מכן שמדובה בארבעה מינים של חיידקים: Bacteroides gracilis, Desulfovibrio gracilis, Curvibacter gracilis, Marichromatium gracile.[3]
בשנות ה-30 של המאה ה-20, פרסם האנולוגיה הצרפתי פסקל ריברו-גיון[4][5] מספר מאמרים שהציגו את היתרונות של פעילות מיקרוביאלית זו ביין.[2] במהלך שנות ה-50, התקדמות בחקר התהליכים האזימתיים אפשרה לאונולוגים להבין טוב יותר את התהליכים הכימיים העומדים בבסיס התסיסה המאלולקטית. אמיל פינו קידם את ההבנה האקולוגית של התהליך, ובמהרה עלתה הזמינות לשימוש בתרביות מבוקרות של חיידקי חומצה לקטית מועילים על ידי ייננים.[3]
בתחילת המאה ה-21, הוכח כי זנים מסוימים של O. oeni משתמשים באצטאלדהיד על ידי פירוקו לאתנול או חומצת חומץ. תהליך זה עשוי לסייע ביינות עם רמות גבוהות מאוד של אצטאלדהיד, כגון, יינות אדומים, אך עלול לערער את יציבות צבע היין, כתוצאה מתגובת האצטאלדהיד עם האנתוציאנינים האחראים ליצירת פיגמנטים פולימריים[6] התורמים צבע היין.[7]


מיקרוביולוגיה של התסיסה המאלולקטית
לכל חיידקי חומצת החלב (LAB) המעורבים בייצור יין, בין אם כתורמים לתסיסה המאלולקטית ובין כמקור לתופעות ארגנולפטיות בלתי רצויות, יש את היכולת לייצר חומצת חלב באמצעות שימוש בסוכר ושימוש בחומצה המאלית כמקור לאנרגיה. מינים שונים נבדלים באופן שבו הם מפרקים את הסוכרים גלוקוז ופרוקטוז, כמו גם פנטוזה הבלתי ניתנת לתסיסה על ידי שמרי היין. מיני חיידקים מסוימים משתמשים בסוכרים דרך מסלול הומו-פרמנטטיבי, שהתוצר הסופי שלו הוא לקטט, בעוד שאחרים משתמשים במסלולים הטרו-פרמנטטיביים שבו נוצרים תוצרים סופיים מרובים: פחמן דו-חמצני, אתנול ואצטט. חיידקי חומצה לקטית הופכים את החומצה המאלית לחומצה לקטית כמקור לאנרגיה על ידי תהליך של כמיאוסמוזה (אנ') שבה נעשה שימוש במפל האלקטרוכימי הנגרם על ידי הבדלי רמות החומציות בין פנים התא לסביבה. מפל זה משמש לייצור ATP. הצורה של L-מלאט הנפוצה ביותר ב-pH נמוך של יין היא המונואניונית הטעונה שלילית. החיידקים מעבירים את האניון הזה מהיין לרמת pH גבוהה יותר לקרום התא הגורם למטען שלילי נטו היוצר פוטנציאל חשמלי. הדה-קרבוקסילציה של מאלאט לחומצה L-לקטית משחררת לא רק פחמן דו-חמצני אלא גם צורכת פרוטון, אשר יוצר מפל של ה-pH בין התא לבין הסביבה.[7]
דרישות מטבוליות של חיידקי התסיסה המאלולקטית
חיידקי חומצה לקטית, הם אורגניזמים בררנים (אנ') שאינם יכולים להרכיב לעצמם את כל מרכיבי התזונה המורכבים העומדים בדרישות התזונתיות שלהם. על מנת שחיידקים אלה יגדלו וישלימו תהליכים של תסיסה מאלולקטית, היין חייב לספק את צרכיו התזונתיים. שמרי יין דורשים מקור פחמן למטבוליזם של אנרגיה, שמקורה לרוב הוא כלל סוכרים והחומצה המאלית, מקור חנקן, כגון חומצות אמינו ופורינים, ליצירת חלבונים, ויטמינים שונים, כגון ניאצין, ריבופלבין ותיאמין, ומינרלים המיועדים לסייע לסינתזה של אנזימים ורכיבים תאיים נוספים אחרים.[8]
המקור לחומרים מזינים אלה נמצא לעיתים קרובות בתירוש הענבים עצמו, אם כי הוספת תרביות חיידקים מאלולקטיים MLF המבוצעת במקביל לתסיסה אלכוהולית מסכנים את השמרים בתהליך תחרותי על מקורות החומרים האלה. לקראת סוף התסיסה, בעוד שרוב משאבי תירוש הענבים המקוריים נצרכו, פירוק תאי שמרים מתים ("המשקעים") יכול להיות מקור לחומרים מזינים מסוימים, במיוחד חומצות אמינו. בנוסף, אפילו יינות "יבשים" שתססו עד יובש עדיין מכילים סוכרים פנטוזיים בלתי ניתנים לתסיסה (אנ'), כגון, אראבינוז, ריבוז וקסילוז, המשמשים הן חיידקים רצויים והן חיידקים מקלקלים כמקור לפחמן ואנרגיה. יצרני תרביות LAB מתורבתים מציעים בדרך כלל תוספי תזונה ייחודיים, המשמשים כתוסף תזונה. בניגוד לשמרי היין, חיידקי חומצה לקטית אינם יכולים להשתמש בתוסף דיאמוניום פוספט כמקור לחנקן.[7]
גם חמצן יכול להיחשב כחומר מזין עבור חיידקי LAB כמו גם לשמרים, אך בכמות קטנה מאוד ורק עבור חיידקים מיקרואארופיליים, כמו O. oeni. לא קיימות ראיות המצביעות על כך שתסיסה מאלולקטית מתקיימת בצורה טובה יותר בתנאים אירוביים מאשר בתנאים אנאירוביים מלאים, וכמויות עודפות של חמצן עלולות לעכב את צמיחתם של חיידקי LAB מפני שימוש מועדף בתנאים אווירניים על ידי חיידקים מתחרים, כמו חיידקי התסיסה של חומצת החומץ האצטובקטר, הגורמים לקילקול היין (אנ').[9]
החיידק Oenococcus oeni המועדף על ידי ייננים לביצוע התסיסה המאלולקטית מצוי באופן טבעי על פני קליפת הענבים, ברמות נמוכות מאוד. פירות מעופשים ופגומים עלולים לשאת פלורה מגוונת של חיידקים שעלולים להתערב בתהליך התסיסה ולגרום ליצירת חומרי לוואי בלתי רצויים. חיידקי ה-LAB הנמצאים לרוב על זגי ענבים נקיים ובריאים שייכים לשני הסוגים: לקטובצילוס, ופדיוקוקוס. התירוש שלאחר ריסוק הענבים מכיל אוכלוסיות מתחת לרמות של 10³ יחידות יוצרות מושבות 9CFU)/מ"ל, ומכילות תערובת של מיני חיידיקים L. rhamnosus, L. casei,L. hilgardii ו-L. plantarum וגם Oenococcus oeni. בתירוש שלא קיבל מנה מוקדמת של גופרית דו-חמצנית כדי "לדכא" את אוכלוסיות-הבר של חיידקי LAB. חיידקי הפלורה של החיידקים הטבעית מתחרים אלה באלה ובשמרי היין, על חומרים מזינים בשלב מוקדם של התסיסה.[9] לכן, הייננים מעדיפים להוסיף סטארטרים מתורבתים של חיידקי LAB במטרה ליצור תרבית מועדפת של חיידקי הסטארטר ולא לאפשר לזני בר להשתלט על אוכלוסיית החיידקים.. התסיסה המאלולקטית מתבצעת על ידי שלושה סוגים של חיידקים. ייננים מונעים באופן פעיל המרה מאלולקטית כאשר היא אינה רצויה, למשל בייצור יין מזני ענבים לבנים פירותיים ופרחוניים כמו ריזלינג וגווירצטרמינר, כדי לשמור על פרופיל חמצמץ או חמצמץ יותר ביין המוגמר.[10][8]
סטארטרים של תסיסה מאלולקטית
-
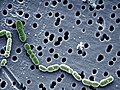 צילום מיקרוסקופ אור של החיידק Oenococcus oeni
צילום מיקרוסקופ אור של החיידק Oenococcus oeni -
 צילום מיקרקרוסקופ אלקטרונים Lactococcus lactis
צילום מיקרקרוסקופ אלקטרונים Lactococcus lactis


בייצור יין מבוקר, התסיסה המאלולקטית מתחילה בדרך כלל על ידי זריעה של סטארטרים מסחריים המכילים את החיידקים הרצויים. במרבית התעשייה משתמשים בתרביות מבוקרות של חיידק Oenococcus oeni המשמש כסטארטר של תסיסה. הזריעה של חיידקי המטרה מונעת מזני חיידקים בלתי רצויים לייצר טעמי "לוואי". ייננים מונעים באופן פעיל המרה מאלולקטית בלתי רצויה בתירוש של זני ענבים לבנים פירותיים ופרחוניים, כגון, זני הענבים ריזלינג וגווירצטרמינר, במטרה לשמור על הטעם החמצמץ של היין.[10][8]
לסוג Oenococcus משתייך מין יחידי, Oenococcus oeni כסטארטר עיקרי בתעשיית היין,[11] חיידק אל-אווירני, פקולטטיבי, גראם- חיובי, המשתמש בחלק מהחמצן לנשימה תאית-אווירנית, אך בדרך כלל מייצר אנרגיה תאית באמצעות תסיסה הטרו-פרמנטטיבית (אנ') של גלוקוזה.[12] בתסיסה זו נוצרים תוצרים סופיים מרובים: החומצה הלקטית-D ופחמן דו-חמצני, המיוצרים בכמויות שוות ערך לאתנול או לאצטט. נוצרת סביבה מחזרת בסמוך לסיום התסיסה האלכוהולית, התוצר הסופי השלישי הוא בדרך כלל אתנול, בעוד שבתנאים אווירניים קלים, למשל, בתחילת תסיסת האלכוהול או בחבית ללא מכסה הקרויה Ullage (wine) החיידקים נוטים לייצר אצטט.[9] זני Oenococcus oeni מסוימים יכולים להשתמש בפרוקטוז כמקור לאנרגיה, בעוד שזנים רבים אחרים יכולים לפרק את חומצת האמינו ארגינין המצויה ביין, שמקורה משמרים בסיום התסיסה הכוהלית, שעברו תהליך של אוטוליזה (אנ') של תאים מתים והפיכתה לאמוניה.[7] בנוסף לחד-סוכרים בעלי שישה פחמנים, ההקסוזות (אנ') גלוקוז ופרוקטוז המצויות ביין, רוב זני O. oeni יכולים להשתמש גם בשאריות של חד-סוכרים בעלי חמישה פחמנים, הפנטוזות (אנ') שנותרו מתסיסת השמרים, ה־L-אראבינוז וריבוזה. רק כ-45% מזני O. oeni יכולים להתסיס סוכרוז שהוא דו-סוכר, המורכב מלגלוקוז ופרוקטוז, המפורק בתהליך התסיסה על ידי שמרים.[7]
זנים מסוימים של O. oeni יכולים לסנתז אלכוהולים גבוהים יותר, התורמים לארומה של היין טעמים פירותיים. זנים מסוימים של החיידק מכילים אנזימי בטא-גלוקוזידאזה (אנ') המפרקים מונוגלוקוזידים (אנ') שהן תרכובות ארומטיות המחוברות למולקולת סוכר. כאשר רכיב הסוכר מתפרק, שאר התרכובת מתנדפת ומגיעה לידי ביטוי בארומה של היין.[7]
לקטובצילים
כל הלקטובצילים המעורבים בייצור יין הם גראם-חיוביים, מיקרואארופיליים, לרובם יש מחסור באנזים קטלאזה המגן מפני עקה כתוצאה מהיווצרות של פראוקסידים.[7] קבוצת הלקטובצילים מכילה מינים הטרו-פרמנטטיביים והומו-פרמנטטיביים.[13] רוב הלקטובצילים אינם רצויים בתהליך ייצור היין בשל הפוטנציאל ליוצרות רמות גבוהות של חומצות נדיפות, ריחות לוואי, והיווצרות יין עכור (אנ'), גזים ומשקעים (אנ') בבקבוק, כאשר היין לא עבר סינון. לחיידקים אלה יש גם פוטנציאל לייצר כמויות עודפות של חומצה לקטית, המשפיעה מאוד על הטעם של היין. מינים מסוימים גורמים לתסיסה איטית ומתמשכת, בעוד שמינים אחרים, כמו Fructilactobacillus fructivorans, ידועים כבעלי יכולת ליצור Lactococcus lactisצמיחה דמוית תפטיר לבן דמוי צמר גפן המצטבר על פני השטח. הצמיחה מכונה "עובש פרזנו" על שם אזור היין בו התגלה.[9]
פדיוקוקוס
ארבעה מינים מהסוג Pediococcus בודדו ביינות ובתירוש ענבים ,P. inopinatus, P. pentosaceus, P. parvulus ו-P. damnosus. שני המינים האחרונים הם הנפוצים ביותר ביין. זהו חיידק גראם-חיוביים, חלקם מיקרו-אארופיליים בעוד שאחרים משתמשים בעיקר בנשימה אירובית. לעיתים קרובות צומחים בזוגות או טטרדות. הם הומופרמנטרים, המפרקים גלוקוז לתערובת רצמית של L- ו-D-לקטט בתהליך של גליקוליזה.[8] בהיעדר גלוקוז, מינים מסוימים, כמו P. pentosaceus, משתמשים בגליצרול, ומפרקים אותו לפירובט, אשר הופך לדיאצטיל, אצט (אנ') ותרכובות אחרות, המקנות ליין מאפיינים אורגנולפטיים בלתי רצויים.[7] רוב מיני החיידקים השייכים לסוג פדיוקוקוס אינם רצויים בייצור יין בשל האפשרות להגיע לרמות הגבוהות של דיאצטיל ויצור מוגבר של אמינים ביוגניים (אנ') ביין.[14] תוצרים אלה הם בעלי פוטנציאל לגרום לכאבי ראש מיין אדום (אנ'). מינים רבים של פדיוקוקוס עלולים לייצר ריחות לוואי בלתי רצויים או פגמים אחרים, כגון, "כתם אקרוליין" בעל טעם מר, הנובע מפירוק גליצרול לאקרוליין, המגיב עם תרכובות פנוליות ביין (אנ') וליצירת תרכובת בעלת טעם מר.[9]
לחלק ממיני הלקטובצילים והפדיוקוקוס (בפרט P. damnosus ו-P. pentosaceus) יש פוטנציאל לסנתז פוליסכרידים המוסיפים צמיגות שמנונית ליין. במקרה של לקטובצילוס, חלק מהסכרידים הללו, הגלוקנים (אנ') המסונתזים מגלוקוז המצויה ביין ברמות נמוכות של 50–100. מ"ג/ליטר, מסונתזים בחביות. יצירת הגלוקנים במכלי התסיסה גורמת לקושי משמעותי בסינון היין לקראת תהליך המילוי[15] הגלוקנים פוגעים ביינות "יבשים" וגורמים ל"אפלוליות". לעיתים קרובות המראה העכור נצפה מספר חודשים לאחר הבקבוק. יינות עם רמות pH מעל 3.5 ורמות נמוכות של גופרית דו-חמצנית נמצאים בסיכון הגבוה ביותר לפתח תקלה זו.[9]
תכונות אורגנולפטיות בתסיסה המאלולקטית
שלב ההבשלה של הענבים הוא נקודת ציון חשובה עבור הכורמים והייננים, שכן הוא מסמן את התקרבות עונת הבציר. תזמון ההבשלה ואחידותו באשכולות יכולים להשפיע רבות על איכות הענבים ובסופו של דבר על איכות היין. ענבים שנקטפו באקלים קריר יותר, בדרך כלל בעלי תכולה הגבוהה יותר של חומצה מאלית וחווים את השינויים הדרמטיים ביותר של ירידה ברמות החומציות הניתנת לטיטרציה וה-pH לאחר תסיסה מאלולקטית.[2] שלב זה מאופיין בשינויים רבים, כגון, שינויי צבע מירוק לגוונים של אדום או סגול, בעוד שענבים המיועדים ליין לבן הופכים מירוקים לירוקים-צהבהבים או זהובים ושקופים יותר. שינוי צבע זה נגרם מהצטברות פיגמנטים כמו אנתוציאנינים בענבים אדומים וקרוטנואידים בענבים לבנים. הענבים הופכים רכים יותר למגע, חלה עלייה ברמות הגלוקוז והפרוקטוז התורם לרמת האלכוהול וירידה ברמת החומציות, ועל ידי כך איזון הטעמים ומתחילים להתפתח רכיבי הטעם והארומה האופייניים לזן הענבים המסוים. תסיסה מאלולקטית נוטה ליצור שינויים בתכונות האורגנולפטיות של היין המתבטאת בתחושה עגולה ומלאה יותר בפה (אנ'). חומצה מאלית מקושרת בדרך כלל לטעם של תפוחים ירוקים, בעוד שחומצה לקטית עשירה יותר וטעמה חמאתי. ענבים המיוצרים באזורים קרירים נוטים להיות בעלי חומציות גבוהה, שחלק ניכר ממנה נובע מתרומתה של החומצה המאלית. תסיסה מאלולקטית בדרך כלל משפרת את התכונה המכונה "הגוף של היין" Body of wine[16] שהיא פועל יוצא של צמיגות היין, ואת עמידות הניחוח של היין, ומייצרת יינות בעלי רכות רבה יותר בתחושת החך. ייננים רבים סבורים כי ניתן להשיג שילוב טוב יותר בין מין הפרי לבין החבית היין העשויה מעץ אלון, אם תסיסה מאלולקטית מתרחשת במהלך שהיית היין בחביות אלה.[2]
יין שעובר תסיסה מאלולקטית יהיה עכור כתוצאה מצמיחת חיידקים. לפעמים מתפתח ריח דמויי פופקורן בחמאה כתוצאה מיצור של דיאצטיל.(אנ')[17] תחילתה של תסיסה מאלולקטית בבקבוק תיראה לצרכן כאילו הוא עדיין בתהליך תסיסה.[18] התפקיד העיקרי של תסיסה מלולקטית הוא הורדת החומציות.[2] זה יכול גם להשפיע על ההיבטים החושיים של יין, לגרום לתחושת חלל הפה (אנ') להיות חלקה יותר ולהוסיף מורכבות לטעם ולארומה שלו. תסיסה מאלולקטית מקטינה את חומציות היין על ידי המרת חומצה מאלית המסוגלת לתרום שני קטיונים של מימן ולכן נחשבת לחומצה "קשה" יותר בהשוואה לחומצה לקטית המסוגלת לתרום קטיון מימן אחד, ונחשבת ל"רכה" יותר. השוני המבני בין חומצות מאלית לבין חומצה לקטית מובילה להפחתה של 1 עד 3 גרם/ליטר בערך החומצה הניתנת לטיטרציה (אנ') (TA) ביין, ולעלייה משמעותית של 0.3 יחידות בערך ה־pH.[8] חומצה מאלית קיימת בענבים לאורך כל עונת הגידול, ומגיעה לשיאה בתקופת ההבשלה, (אנ') שבו חלים השינויים בתחושת הפה קשורים לעלייה ב-pH, אך ייתכן שהוא נובע גם מייצור פוליאולים (אנ') ובמיוחד אלכוהולי הסוכר אריתריטול וגליצרול[7] גורם נוסף שעשוי לשפר את תחושת הפה של יינות שעברו תסיסה מאלולקטית הוא נוכחותו של אתיל לקטט (אנ') עד רמות של 110. מ"ג/ליטר בתום התסיסה המאלולקטית.[8]
התוצרים העיקריים של תסיסה מאלולקטית הם חומצה לקטית, דיאצטיל, חומצת חומץ, אצטואין (אנ') ואסטרים שונים. הכמות והאופי המדויק של מוצרים אלה תלויים במין/זן של חיידקי ה-LAB המבצע את התסיסה המאלולקטית ובתכונות הכימיות המשפיעות על אותו יין : pH, חומרים מזינים זמינים, רמות חמצן וכו').[19]
זנים מסוימים של O. oeni יכולים לסנתז אלכוהולים גבוהים יותר שיכולים לתרום לתווים פירותיים בארומה של היין. זנים מסוימים של החיידק מכילים אנזימי בטה-גלוקוזידזה (אנ')שיכולים לפרק מונוגלוקוזידים (אנ'), שהם תרכובות ארומטיות הקשורות למולקולת סוכר. כאשר רכיב הסוכר מתפרק, שאר התרכובת מתנדפת, וניתנת לחישה בארומה של היין.[7]
דיאצטיל
דיאצטיל (או 2,3-בוטאנדיון) הוא התרכובת המקושרת לארומות ה"חמאתיות" של יין שרדונה, אך הוא יכול להשפיע על כל יין שעבר תסיסה מאלולקטית. בבדיקהמעבדית לגילוי סף גילוי הריח של דיאצטיל נמצא 0.2 מ"ג/ליטר ביינות לבנים ו-2.8 מ"ג/ליטר ביינות אדומים, הוא יכול להיתפס כמעט חמאתי או "אגוזי" בריכוזים 5–7 מ"ג/ליטרומעלים ריחות ארומטיים אחרים.[18][20]
דיאצטיל נוצר בתהליך מטבולי של סוכר או של חומצת לימון.[21] בעוד שחומצת לימון קיימת באופן טבעי ברמה נמוכה בענבים, ורובה הוא תוצאה של הוספה מכוונת של היינן,[9] כאשר חומצות מאלית וחומצת לימון נוכחות, קצב ניצול הסוכר ויצירת הדיאצטיל מהירה יותר בתסיסה המאלולקטית מאשר חומצת חומצת הלימון, והוא מושפע מזן החיידקים הספציפי כשרוב זני
O. oeni מייצרים פחות דיאצטיל מאשר מיני לקטובצילוס ופדיוקוקוס. כמו כן, קצב יצירת הדיאצטיל מושפע מפוטנציאל החיזור של היין.[22][8] דיאצטיל נוצר ברמות גבוהות יותר ביינות עם רמות pH נמוכות מתחת ל-3.5. ברמות מתחת ל-3.2, רוב זני LAB הרצויים לתסיסה המאלולקטית מעוכבים ומפסיקים להתרבות. לתסיסות מאלולקטיות "פראיות" עם זני-בר יש יכולת לייצר יותר דיאצטיל מאשר תסיסות מבוקרות, עקב אוכלוסיות ראשוניות נמוכות יותר בשלב הלג (אנ') Lag phase, כאשר לתסיסות מבוקרות יש בדרך כלל ריכוז התחלתי של 106 CFU/מל.[7] כאשר תהליך התסיסה המאלולקטית מתרחש זמן ארוך אחרי התסיסה האלכוהולית, קיימת נטייה לייצור רמות גבוהות יותר של דיאצטיל.[19] יצרני יין שרדונה המעוניינים לייצר יין "חמאתי" בעל ריכוז גבוה של דיאצטיל, יבצעו לעיתים קרובות תסיסה מאוחרת בחבית לאחר התסיסה הראשונית, המאפשר ליין לשהות מספר שבועות או אפילו חודשים בתנאי חיזור המקדמים יצירת דיאצטיל.[9][23]
ביינות המכילים רמות גבוהות מאוד של דיאצטיל, נוהגים להוסיף גופרית דו-חמצנית המתקשרת לדיאצטיל ומורידה את הרמה ב-30 עד 60%. גופרית דו-חמצנית שמוספת בשלב מוקדם יותר בתהליך התסיסה המאלולקטית מגבילה את ייצור הדיאצטיל, על ידי עיכוב החיידקים והגבלת פעילותם במלואה.[18] התהליך הפיך ולאחר מספר שבועות של יישון היין (אנ') בבקבוק או במכל, חלה עליה ברמות הדיאצטיל.
- מבנים סכמטיים של כהלים נדיפים שנוצרים בתסיסה מאלולקטית
-
 דיאצטיל
דיאצטיל -
 אצטואין
אצטואין -
 גליצרול
גליצרול -
 בוטילן גליקול
בוטילן גליקול



אצטואין, בוטילן גליקול וגליצרול
בעת התסיסה המאלולקטית עשויים להיווצר גם אצטואין (אנ'), חומר נדיף בעל ריח המעניק ליין תכונות אורגנולפטיות אופייניות,[24][25] וכן כוהלים בעלי ריח אופיינים, הבוטילן גליקול,[8] וגליצרול[26] התורמים אף הם ריחות אופייניים ליין.
מניעת תסיסה מאלולקטית
ביינות קלים, פירותיים ובעלי חומציות נמוכה המיוצרים באקלים חם, תסיסה מאלולקטית אינה רצויה. ייננים יכולים לנקוט במספר צעדים כדי למנוע את התרחשותה של תסיסה מאלולקטית בלתי רצויה.[10][3]
- השרייה מוגבלת של הענבים למטרת ריכוך, סחיטה מוגבלת, כבישה (אנ') מוקדמת, ואיסוף מוקדם כדי להגביל את זמן המגע של הפלורה המיקרוביאלית עם מקורות מזון לחיידקים.
- הקפדה על רמות גופרית דו-חמצנית של לפחות 25 מ"ג/ליטר SO2 "חופשי", בהתאם לרמת החומציות של היין. במקרים מסוימים, ייתכן שיהיה צורך להוסיף 50–100 מ"ג/ליטר של SO2
- שמירה על רמות pH מתחת ל-3.3
- שמירת היין בקירור בטמפרטורות שבין 10 ל-14 מעלות צלזיוס
- סינון היין בתהליך הביקבוק דרך מסנן ממברנלי, בעל נקבים בקוטר של עד 0.45 מיקרון, המונע חדירת חיידקים לבקבוק.
- שימוש במעכבים כימיים וביולוגיים כגון ליזוזים, ניסין, (אנ') דימטיל דיקרבונט (אנ') ברמות עד 200 מ"ג/ליטר וחומצה פומרית. חלקם של החומרים האלה מוגבלים לשימוש במדינות ייצור יין מחוץ לארצות הברית (אנ').
- טיפול בחומרי עידון, כגון בנטוניט, וייצוב קר של היין במשך זמנים שונים מסירים חומרים מזינים עבור חיידקי LAB, ובכך עיכוב תסיסה מאלולקטית.
תקלות בתסיסה מאלולקטית
- התקלה הנפוצה ביותר הקשורה לתסיסה מאלולקטית מתרחשת ביין שנועד להיות חומצי ופירותי (כגון ריזלינג)
- היצרן שער שהיין עבר תסיסה מאלולקטית בחביות, אולם התברר שהתסיסה המאלולקטית התחילה למעשה בבקבוק. התוצאה של תסיסה "בתוך הבקבוק" היא לרוב יין גזי עם עכירות דמוית ערפל. מחייב שיפור תנאי התברואה והבקרה על חיידקי החומצה הלקטית ביקב.
- ייצור יין ל"לא יציב". שינוי ה-pH משפיע על מסיסות החלבונים ביין. עובדה זו מחיבת בדיקות עוקבות של יציבות היין בחום לאחר סיום התסיסה המאלולקטית.[8]
- ענבי יין הנגועים בריקבון על ידי העובש בוטריטיס (אנ') נוטים להיות בעלי רמות גבוהות יותר של גליצרול, המתפרק על ידי חיידקי החומצה הלקטית לאקרוליין. בענבי יין אדומים בעלי תכולת פנולים גבוהה עלולים לפתח יינות בעלי טעם מר, בעקבות האינטראקציה בין האקרוליין והפנולים האלה.[27][28]
קישורים חיצוניים
הערות שוליים
- ↑ Tom Mansell "Buttery bacteria: Malolactic fermentation and you (אורכב 06.04.2016 בארכיון Wayback Machine)" Palate Press. 10 November 2009
- ^ 2.0 2.1 2.2 2.3 2.4 2.5 J. Robinson (ed) "The Oxford Companion to Wine" Third Edition pgs 422 & 508 Oxford University Press 2006 מסת"ב 0-19-860990-6
- ^ 3.0 3.1 3.2 3.3 Sibylle Krieger "The History of Malolactic Bacteria in Wine (אורכב 15.09.2012 בארכיון Wayback Machine) pgs 15-21. Accessed: 14 May 2013
- ↑ Suzanne Mustacich, [Wine Spectator Bordeaux Enologist Pascal Ribéreau-Gayon Dies], https://www.winespectator.com/articles/bordeaux-enologist-pascal-ribereau-gayon-dies-45102, May 2011
- ↑ Further understanding of phenolic composition of grapes and wine - Wine History, winehistory.com.au
- ↑ Joana Oliveira, Victor de Freitas, Nuno Mateus, Chapter 14 - Polymeric Pigments in Red Wines, Academic Press, 2019-01-01, עמ' 207–218, מסת"ב 978-0-12-814399-5
- ^ 7.00 7.01 7.02 7.03 7.04 7.05 7.06 7.07 7.08 7.09 7.10 K. Fugelsang, C. Edwards Wine Microbiology Second Edition pgs 29-44, 88-91, 130-135, 168-179 Springer Science and Business Media, New York (2010) מסת"ב 0387333495
- ^ 8.0 8.1 8.2 8.3 8.4 8.5 8.6 8.7 8.8 B. Zoecklein, K. Fugelsang, B. Gump, F. Nury Wine Analysis and Production pgs 160-165, 292-302 & 434-447 Kluwer Academic Publishers, New York (1999) מסת"ב 0834217015
- ^ 9.0 9.1 9.2 9.3 9.4 9.5 9.6 9.7 R. Boulton, V. Singleton, L. Bisson, R. Kunkee Principles and Practices of Winemaking pgs 244-273 & 369-374 Springer 1996 New York מסת"ב 978-1-4419-5190-8
- ^ 10.0 10.1 10.2 Dr. Yair Margalit, Winery Technology & Operations A Handbook for Small Wineries pgs 75-78, 103 & 183-184 The Wine Appreciation Guild (1996) מסת"ב 0-932664-66-0
- ↑ Antonella Costantini, Francesca Doria, Enrico Vaudano, Maria Carla Cravero, Emilia Garcia-Moruno, Selection of Oenococcus oeni as starter cultures to induce malolactic fermentation in Nebbiolo wine, WORLD SCIENTIFIC, 2010-12, עמ' 431–435 doi: 10.1142/9789814322119_0093
- ↑ Ronald S. Jackson, 7 - Fermentation, San Diego: Academic Press, 2014-01-01, Food Science and Technology, עמ' 427–534, מסת"ב 978-0-12-381468-5
- ↑ (cals.cornell.edu), [chrome-extension://efaidnbmnnnibpcajpcglclefindmkaj/https://cals.cornell.edu/sites/default/files/2023-04/lactic-acid-bacteria-v1.pdf Lactic Acid Bacteria – Homofermentative and Heterofermentative], (cals.cornell.edu)
- ↑ M.H.Silla Santos, Biogenic amines: their importance in foods, International Journal of Food Microbiology 29, 1996-04, עמ' 213–231 doi: 10.1016/0168-1605(95)00032-1
- ↑ A Nightmare on Filtration Street, Laffort, 2024-10-30
- ↑ words: VinePair Staff, Wine Body Guide - Light, Medium & Heavy Body!, VinePair
- ↑ Managing Diacetyl Production During MLF, scottlab.com
- ^ 18.0 18.1 18.2 John Hudelson "Wine Faults-Causes, Effects, Cures" pgs 46-53, The Wine Appreciation Guild (2011) מסת"ב 9781934259634
- ^ 19.0 19.1 Jean Jacobson "Introduction to Wine Laboratory Practices and Procedures" pgs 188-191, Springer Science and Business Media, New York (2010) מסת"ב 978-1-4419-3732-2
- ↑ Martineau, B., Acree, T.E. and Henick-Kling, T "Effect of wine type on the detection threshold for diacetyl" Food Research International. Volume 28, Issue 2, 1995, Pages 139–143
- ↑ Shimazu, Y., Uehara, M., and Watanbe, M. "Transformation of Citric Acid to Acetic Acid, Acetoin and Diacetyl by Wine Making Lactic Acid Bacteria" Agricultural and Biological Chemistry 49(7), 2147-2157, 1985
- ↑ Jan Clair Nielsen and Marianne Richelieu "Control of Flavor Development in Wine during and after Malolactic Fermentation by Oenococcus oeni" Applied and Environmental Microbiology. February 1999 vol. 65 no. 2 740-745
- ↑ Rotter, Ben. "Sur lie and bâtonnage (lees contact and stirring)". Improved winemaking, 2008. Retrieved 12-Feb-2016.
- ↑ Zijun Xiao, Jian R. Lu, Strategies for enhancing fermentative production of acetoin: A review, Biotechnology Advances 32, 2014-03-01, עמ' 492–503 doi: 10.1016/j.biotechadv.2014.01.002
- ↑ Graham H. Fleet, Wine Microbiology and Biotechnology, CRC Press, 1993-01-01, מסת"ב 978-0-415-27850-8
- ↑ Ionel Popescu-Mitroi, The study of glycerol metabolism in the malolactic fermentation of red wines, ROMANIAN BIOTECHNOLOGICAL LETTERS 19(1):9019 - 9027, 2014
- ↑ Rolene Bauer, Donald A. Cowan, Andrew Crouch, Acrolein in wine: importance of 3-hydroxypropionaldehyde and derivatives in production and detection, Journal of Agricultural and Food Chemistry 58, 2010-03-24, עמ' 3243–3250 doi: 10.1021/jf9041112
- ↑ María Perez-Jiménez, Adelaida Esteban-Fernández, Carolina Muñoz-González, María Angeles Pozo-Bayón, Interactions among Odorants, Phenolic Compounds, and Oral Components and Their Effects on Wine Aroma Volatility, Molecules 25, 2020-04-08, עמ' 1701 doi: 10.3390/molecules25071701

תסיסה מאלולקטית41749225Q654005